
雄蕊
雄蕊(雄蘂、雄ずい [ゆうずい]、雄しべ [おしべ]; 英: stamen)[1]は、被子植物の花において花粉を形成する構造であり、ふつう花被(萼や花冠)の内側、雌蕊(雌しべ)の外側に存在する。雄蕊はふつう花粉を形成する葯(やく; anther)[2]と、これを支える花糸(かし; filament)[3]からなる(下図1)。ふつう葯は2個の半葯からなり、それぞれの半葯は2個の花粉嚢(小胞子嚢、葯室)を含む。花粉嚢内で形成された花粉は(別の花または同じ花の)雌蕊の柱頭に付着し(受粉)、花粉は花粉管を伸ばして雌蕊内の胚珠に精細胞を送り込み、胚珠内の卵細胞と受精する。1個の花の雄蕊は、まとめて、雄蕊群(雄ずい群、雄しべ群; androecium, pl. androecia)とよばれる[1][4][5]。



雄蕊は、被子植物における雄性胞子葉(小胞子葉; 雄性胞子嚢をつけた葉的構造)に相当する[6]。裸子植物の花粉をつくる構造(小胞子葉)も雄蕊(雄ずい、雄しべ)とよばれることがあるが、被子植物の雄蕊との厳密な相同性は必ずしも明らかではないため、生物学的にはこれを雄蕊とよばないことが多い[5]。
構成
[編集]
雄蕊(雄ずい、雄しべ)は被子植物の雄性生殖器官であり、ふつう葯とそれを支える花糸からなる[4][5][7][8][9][10](図2)。葯の花粉嚢(雄性胞子嚢、小胞子嚢、葯室)の中で、減数分裂によって花粉が形成される。
花糸
[編集]雄蕊において、葯を支える構造は花糸とよばれる[4][5][8][9][10]。花糸はふつう細長い軸状であるが(下図3a, b)、扁平な葉状であるものもいる(下図3c–e)。このような花糸をもつ雄蕊は、葉状雄蕊(葉状雄ずい、葉的雄ずい、葉状雄しべ; laminar stamen)とよばれる[11]。スイレンでは、花の外側から内側に向かって葉状(花弁状)の花糸から糸状の花糸まで連続的な移行が見られる[4][9](下図3c)。葉状雄蕊は被子植物の中で初期に分岐した植物群にしばしば見られ(下図3d, e)、デゲネリア科やバンレイシ科、ヒマンタンドラ科では背軸側(裏側)に、モクレン属やアウストロバイレヤ科では向軸側(表側)に花粉嚢がついている[9][11][12]。このような雄蕊では、しばしば花糸と葯が明瞭に分化していない[4][8]。このような葉状雄蕊は原始的な状態を示すものとされることもあるが[11][12]、被子植物の初期分岐群の中にも糸状の花糸をもつものがおり(下図3b)、花糸の形態に関する進化は独立に何回も起こったと考えられている[11][6]。このような糸状の花糸は、葉の背軸側の遺伝子のみが花糸全体で発現しているために生じると考えられている(葉では、背軸側・向軸側の遺伝子が発現することでその境界が伸長して扁平化する)[6]。
.jpg)


_fragment-2.jpg)

ふつう、花糸には1本の維管束があるが(上図2)、葉状雄蕊では複数の維管束が縦走していることもある[9][11]。一方で、退化的で維管束を欠いていることもある[9]。
花糸には、しばしば腺毛や軟毛などが生じている[4][8](下図4a)。また花糸に蜜腺のような特殊な構造が付随している例(クスノキ属など; 下図4b)や、花糸の基部が翼状に広がっている例(ウメガサソウ属; 下図4c)もある[4]。
明瞭な花糸を欠く雄蕊も存在し、無柄雄蕊(無柄雄ずい、無柄雄しべ; sessile stamen)とよばれる[4]。無柄雄蕊は、ウマノスズクサ属、アマモ属、イバラモ属、クルミ属などに見られる[4](下図4d)。
トウゴマ(トウダイグサ科)の"雄花"では、"花糸"が細かく分枝しており、それぞれの先端に葯をつけている[4](下図4e)。この構造は多数の花糸または雄花が融合したものと考えられている[13]。

_y_ap%C3%A9ndices_nectar%C3%ADferos.jpg)
.JPG)

_-_Flickr_-_Kevin_Thiele.jpg)
葯
[編集]
雄蕊において、花粉を形成する構造的まとまりは葯とよばれる[4][7][2][14]。葯は、ふつう左右にならんだ2個の半葯(はんやく; theca, pl. thecae)[14][15]とその間をつなぐ葯隔(やくかく; connective)[2]からなる(bithecal[16])[4][7][14](図5)。1個の半葯には、ふつう2個の花粉嚢(pollen sac; 小胞子嚢 microsporangium; 葯室 anther locule)が含まれる[4][7][17](図5)。よって、1個の雄蕊はふつう4個の花粉嚢を含む(tetrasporangiate[16])。
上記のように葯はふつうそれぞれ2個の花粉嚢を含む2個の半葯からなるが、ツリフネソウ属の一部は、3個の半葯からなる葯をもつ[4]。またイワブクロ属(オオバコ科)のように、1個の半葯からなる葯をもつものもいる[4](monothecal, bisporangiate[16])。ワサビノキ属(ワサビノキ科)やミジンコウキクサ属(サトイモ科)では、2個の半葯がそれぞれ花粉嚢を1個のみ含む[18]。さらに Arceuthobium(ビャクダン科)は、1個の雄蕊が花粉嚢を1個のみをもつ[18]。
被子植物の花粉嚢は他の陸上植物の雄性胞子嚢よりも大きいことから、複数の雄性胞子嚢が癒合したものではないかとする考えもあるが、発生過程においてそれを支持する証拠は見つかっていない[6]。
葯隔
[編集]葯において、2個の半葯をつなぐ位置にある組織は葯隔とよばれる[4][14](図5, 下図6a)。沿着葯や内着葯(下記参照)をもつものでは、花糸と葯隔が連続的で両者の境界が不明瞭なこともある。
葯隔は特殊な構造をしていることもある。ツクバネソウ(シュロソウ科)やアキカラマツ(キンポウゲ科)では、葯隔が先端側に突出している[4](下図6b)。スミレ属(スミレ科)では、葯隔の先端が褐色膜質の付属体になる[4](下図6c)。また下側の2個の雄蕊の葯隔から突起が生じ、1個の花弁(唇弁)から構成される細長い突出部(距)に入っている(蜜を分泌する)[4](下図6c)。アキギリ属(シソ科)では半葯が上下に離れて配置しており(下側の半葯は花粉形成能を欠く)、その間が長い葯隔でつながっている[4](下図6d)。花に侵入した送粉者が下側の半葯を押すと、てこの原理で上側の半葯が下がって送粉者の背中に花粉をつける(下図6e)。
.jpg)
.jpg)


_sl12.jpg)
葯のつき方
[編集]花糸に対して葯がどのようについているのかには、以下のような多様性がある。
- 側着(沿着、側沿着、adnate)[4][8][19](下図7a)
- 葯の全長が花糸の延長部に接しており、花糸と葯隔が連続的なこともある。最もふつうに見られる。
- 内着(innate)[4](下図7b)
- 葯が花糸の延長部の組織中に埋没しており、葯の組織が突出していない。花糸と葯隔が連続的である。メギ属(メギ科)やサバノオ(キンポウゲ科)などに見られる。
- 底着(basifixed)[4][8][5][20](下図7c)
- 花糸が先端に向かって細くなり、その先端に葯の下端がついている。キンコウカ(キンコウカ科)、カタクリ属(ユリ科)、ユキザサ(キジカクシ科)、ヒサカキ属(モッコク科)などに見られる。
- 丁字着(ていじちゃく; versatile)[4][8][5][20](下図7d)
- 花糸の先端が葯の側面の一点についている。ユリ属(ユリ科)、カモガヤ(イネ科)、ツバキ属(ツバキ科)、マツヨイグサ属(アカバナ科)、オオバコ属(オオバコ科)などに見られる。




葯の向き
[編集]葯の向きは、以下のように多様である[4][21]。動物媒花(昆虫など動物によって花粉媒介される花)において、このような葯の向きは送粉者の行動と密接に関わっており、花において送粉者が行動する方向に向いている[4]。葯では、葉の向軸側と背軸側の遺伝子が発現し、その境界部で花粉嚢が形成されることが報告されており、この向軸側・背軸側遺伝子の発現領域の変化が下記のような葯の向きに関わっていることが示唆されている[6]。
- 外向葯(外開葯、extrorse anther)[4][7][22](下図8a)
- 花の外側に向かって裂開する葯のことであり、花糸の背軸側に位置する。ユリノキ、ウマノスズクサ科、サトイモ科、ヒルムシロ科、アヤメ科、イグサ科、アケビ科、タデ科、ウリ科、イカリソウ、ヤマモモ科などに見られる。
- 側向葯(laterorse anther; 側開葯 equifacial anther)[4][7][23](下図8b)
- 雄蕊の左右に向かって裂開する葯のことであり、花糸の側面に位置する。ヤマグルマ、マンサク科、スズカケノキ科、フサザクラ科、カツラ科、ナデシコ科、ハマミズナ科、ウルシ科、カタバミ科、ヒルガオ科、クマツヅラ科などに見られる。
- 内向葯(内開葯、introrse anther)[4][7][24](下図8c)
- 花の内側に向かって裂開する葯のことであり、花糸の向軸側に位置する。スイレン科、ヒユ科、キョウチクトウ亜科、キク科などに見られる。
ふつう1つの花の中の複数の雄蕊において葯の向きは一定であるが、1つの花の中で葯の向きが異なる雄蕊が共存することもある。クスノキ属などでは3個ずつ3輪の雄蕊があるが(仮雄蕊を除く)、第1、第2輪の雄蕊は内向葯をもち、第3輪の雄蕊は外向葯をもつ[4](下図8d)。

.JPG)

_Mez_-_Flickr_-_Alex_Popovkin,_Bahia,_Brazil_(43).jpg)
葯の裂開
[編集]


葯は裂開して花粉を放出する。この際の裂開様式には、以下のような多様性がある[4]。
- 縦裂(longitudinal dehiscence)[4][25](図9a, 下図10a–c)
- 葯が縦方向に裂開する。個々の花粉嚢が裂開する場合と、半葯の2個の花粉嚢の隔壁が崩壊してその部分で裂開する場合がある。最もふつうに見られる裂開様式である。
- 横裂(transverse dehiscence)[4](右図9b)
- 葯が横方向に裂開する。シュロソウ属(シュロソウ科)、ツリフネソウ属(ツリフネソウ科)、フヨウ属(アオイ科)、ネコノメソウ属(ユキノシタ科)、トウダイグサ属(トウダイグサ科)などに見られる。
- 孔開(poricidal dehiscence, porous dehiscence)[4][26](上図4c, 図9c, 下図10d, e)
- 葯の一部の組織が喪失して孔があく。このような葯は孔開葯 (poricidal anther, porous dehiscent anther) という。ツツジ科の多くやナス属(ナス科)などに見られる。
- 弁開(弁状裂開; valvular dehiscence)[4](上図4b, 図9d, 下図10f)
- 葯の一部がめくれ上がって開口する。このような葯は弁開葯(valvular anther, porous dehiscent anther)という。クスノキ科やメギ科の多くに見られる。






数と配置
[編集]基本的に、1つの花の中の雄蕊(雄ずい、雄しべ)の数や配置は、種によって決まっている。
スイレン属やモクレン属、オモダカ属、キンポウゲ属、ケシ属、バラ属などでは、1個の花が不特定多数の雄蕊をもつ[8](下図11)。スイレン属やモクレン属では、多数の雄蕊がらせん状に配列しており、一般的に、このような状態が被子植物における原始的な状態であると考えられていた[12]。ただし、雄蕊の二次的な不特定多数化は、被子植物の中で何度も独立に起こっていると考えられている[27]。フヨウ属(アオイ科)やツバキ属(ツバキ科)も不特定多数の雄蕊をもつが、これらの雄蕊は互いに合着している(下記参照)。
.jpg)




_(4769245072).jpg)
決まった数の雄蕊をもつ花では、基本的に花被片の基本数(ふつう 3, 4 または 5)と同じ数の雄蕊が、1輪(uniseriate)または内外2輪(biseriate)に輪生している (whorled)[5][8][28]。例えばユリ属やイネ属などでは3個ずつ(下図12a)、マツヨイグサ属などでは4個ずつ(下図12b)、ナデシコ属やカタバミ属などでは5個ずつ(下図12c)の雄蕊が2輪に互い違いに配置している。またドクダミ属やアヤメ属、ススキ属などでは3個(下図12d)、ヤエムグラ属などでは4個(下図12e)、スミレ属やキキョウ属などでは5個(下図12f)の雄蕊が1輪に配置している。ただしこのような基本形から雄蕊数が減少しているものも多く、例えばアブラナ科ではふつう外輪に2個、内輪に4個の雄蕊が存在し、またクワガタソウ属では雄蕊は2個のみ存在する(下図12g)。極度に単純化した花では、雄蕊1個のみからなる例もある(例: トウダイグサ属)。







雄蕊は基本的に花被と雌蕊(雌しべ)の間に存在するが、雌蕊を欠く雄花では、雄蕊が花の中央に位置する。またラカンドニア・スキスマティカ(ラカンドニア科)では、多数の雄蕊が花の中央に位置し、その周囲を多数の雌蕊が取り囲んでいる[29]。
雄蕊の合着
[編集]1つの花の中で、雄蕊どうしが合着(同類合着)、または雄蕊が他の花要素と合着(異類合着)していることがある[4]。
雄蕊の同類合着
[編集]雄蕊の花糸どうしが合着しているもの、葯どうしが合着しているもの、全体が合着しているものがある。
花糸どうしが合着
[編集]葯は分離しているが、少なくとも花糸の一部どうしが合着している雄蕊は、合糸雄蕊(合糸雄ずい、合糸雄しべ; adelphous stamen)とよばれる[4][5]。合糸雄蕊を形成することで、1個の花の中で何組の雄蕊が存在するのかによって、以下のように分けられる。
- 単体雄蕊(単体雄ずい、単体雄しべ; monadelphous stamen)[4][5][8][30]
- 1つの花の中で、すべての雄蕊の花糸が合着して単一の構造になっているもの(下図13a, b)。合着した花糸によって筒(花糸筒 filamental tube)が形成され、その中を雌蕊が貫通している。ツバキ科、アオイ科、マメ科の一部(ネムノキ属、ハギ属、レンリソウ属など)などに見られる。
- 二体雄蕊(二体雄ずい、二体雄しべ、両体雄ずい、両体雄しべ; diadelphous stamen)[4][5][8][31]
- 1つの花の中で、一部の雄蕊の花糸が合着して2組になっているもの(下図13c)。ケマンソウ属(ケシ科)は6本の雄蕊をもつが、3本ずつ花糸が合着して2組に分かれている。マメ科の多く(イワオウギ属、シャジクソウ属、ヤブマメ属など)は10本の雄蕊をもつが、そのうち9本の花糸が合着して樋のような構造を形成し、1本だけが他と離れている。
- 多体雄蕊(多体雄ずい、多体雄しべ; polyadelphous stamen)[4][32]
- 1つの花の中で、一部の雄蕊の花糸が合着して3組以上になっているもの。オトギリソウ属の多くでは雄蕊が3組に分かれており、三体雄蕊(三体雄ずい、三体雄しべ; triadelphous stamen)[4][5][33] とよばれ(下図13d)、トモエソウやシナノキ属では雄蕊が5組に分かれており、五体雄蕊(五体雄ずい、五体雄しべ; pentadelphous stamen)[4][8][34] とよばれる。


.jpg)

葯どうしが合着
[編集]花糸は分離しているが、葯が互いに合着している雄蕊は、集葯雄蕊(集約雄ずい、集約雄しべ; syngenesious stamen; synandrium, pl. synandria; synangium, pl. synangia)とよばれる[4][5][8][35](下図14a–c)。ツリフネソウ科やキク科のほとんどにみられる。
全体が合着
[編集]花糸も葯も全体が合着している雄蕊は、合体雄蕊(合体雄ずい、合体雄しべ; synandreous stamen)とよばれる[4]。ミゾカクシ属(キキョウ科)の一部やアマチャヅル属、アレチウリ属(ウリ科)などでは1つの花の中の全ての雄蕊が合着して合体雄蕊を形成している[4](下図14d, e)。またスズメウリ属やミヤマニガウリ属(ウリ科)では、5本の雄蕊のうち2本ずつ2組が合着して合体雄蕊を形成している[4]。


.jpg)
_2_uni%C3%B3n_de_filamentos_y_anteras,_detalle,_sin_orificio.jpg)
雄蕊の異類合着
[編集]花被との合着
[編集]雄蕊は、しばしばその外側にある花被と合着している。サクラソウ、リンドウ、ムラサキ、シソ、ゴマノハグサ、キツネノマゴ、アカネ、オミナエシ、ヒマワリなど、いわゆる合弁花冠(花弁どうしが合着している花冠)をもつ花では、雄蕊の花糸が花冠に合着していることが多い[4]。このような雄蕊の状態は花冠上生(花冠着生、epipetalous)とよばれる[4][5][36](下図15a, b)。
アヤメやフリージア、グラジオラス、アマドコロ、スズランなど花被が萼と花冠に分化していない植物でも、花被に雄蕊の花糸が合着していることがあり、このような雄蕊の状態は花被上生 (epitepalous) とよばれる[4](下図15c)。
花被を1輪だけもつ植物(このような植物の花は単花被花とよばれる)では、その花被はしばしば萼とよばれる。グミやジンチョウゲでは、花被(萼)に雄蕊の花糸が合着しており、このような雄蕊の状態は萼上生(epsepalous)とよばれる[4](下図15d)。
.jpg)


.jpg)
雌蕊との合着
[編集]植物群によっては、雄蕊が雌蕊(雌しべ)に合着しており、このような雄蕊の状態は雌蕊着生(epigynoecious)とよばれる[4]。センリョウ科では雌蕊の子房に雄蕊が合着しており(下図16a)、ウマノスズクサ科では1輪の雄蕊が雌蕊を取り囲んでこれに合着している(下図16b)。
ラン科やガガイモ類(キョウチクトウ科)では雄蕊と雌蕊の合着が極度に進み、蕊柱(ずいちゅう、column; gynostemium, pl. gynostemia)とよばれる1個の構造となっている[4][37](下図16c, d)。
_Sarcandra_glabra_-%E9%A6%99%E6%B8%AF%E8%A5%BF%E8%B2%A2%E7%8D%85%E5%AD%90%E6%9C%83%E8%87%AA%E7%84%B6%E6%95%99%E8%82%B2%E4%B8%AD%E5%BF%83_Saikung,_Hong_Kong-_(9229827600).jpg)


_Raf.,_Fl._Tellur._2_38_(1837)_(50296155292).jpg)
異形雄蕊
[編集]1つの花の中で、異なる形や大きさの雄蕊が存在する場合、このような雄蕊は異形雄蕊(異形雄ずい、異形雄しべ; heteromorphous stamen)とよばれる[4]。ただし1つの花の中に不特定多数の雄蕊がある場合、連続的に大きさなどの変化が見られることが多いが、これは異形雄蕊とはよばない。また1つの花の中に通常の雄蕊と仮雄蕊(不稔の特殊化した雄しべ)が共存する場合も、異形雄蕊とはよばない。
長さが異なる異形雄蕊
[編集]1つの花の中で、特定のいくつかの雄蕊のみが長いことがある。長い雄蕊の数によって、以下のように類別される。
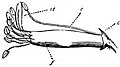
- 二強雄蕊(二強雄ずい、二強雄しべ、二長雄ずい、didynamous stamens)
- 4本ある雄蕊のうち、2本が長いもの[4][5][8](下図17a, b)。シソ科やハマウツボ科、オオバコ科、ハエドクソウ科などに見られる。
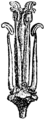
- 四強雄蕊(四強雄ずい、四強雄しべ、四長雄ずい、tetradynamous stamens)
- 6本ある雄蕊のうち、4本が長いもの[4][5][8](下図17c)。アブラナ科に特有であり、2輪あるうちの内輪の4個の雄しべ(花弁と対生する)が長い。
- 五強雄蕊(五強雄ずい、五強雄しべ; pentadynamous stamens)
- 10本ある雄蕊のうち、5本が長いもの[4](下図17d, e)。ミソハギ科やカタバミ科では、2輪あるうちの外輪の5個の雄しべが長いが、フウロソウ属では内輪の5個の雄しべが長い。




_close-up_(9194673524).jpg)
仮雄蕊
[編集]構造としては残っているが、花粉形成能を失っている雄蕊は、仮雄蕊(仮雄ずい [かゆうずい]; 仮雄しべ [かりおしべ]; 化生雄蕊; staminode; staminodium, pl. staminodia)とよばれる[4][5][8][38]。また仮雄蕊において、構造的には残っているが花粉を形成しない葯がある場合、これは仮葯(antherode)とよばれる。1つの花の雄蕊が全て仮雄蕊になっている例と、1つの花の中で決まった雄蕊のみが仮雄蕊になっている例がある。
.jpg)
.jpg)
.jpg)
雌花では、雌蕊に加えてしばしば機能しない雄蕊(仮雄蕊)が存在することがある[4](上図18a–c)。例えばダンコウバイ(クスノキ科)、ヤマノイモ(ヤマノイモ科)、オオヤマフスマ(ナデシコ科)、イタドリ属(タデ科)、カエデ属(ムクロジ科)、カキノキ(カキノキ科)、モチノキ(モチノキ科)などの雌花には、機能する雄蕊は存在せず、仮雄蕊が存在する[4]。
一方、同一の花の中で、通常の雄蕊と仮雄蕊が同居することもある[4](下図19a–c)。ツユクサの花は6個の雄蕊をもつが、黄色く目立つ短い3個の雄蕊は花粉を形成しない仮雄蕊であり、2個の長い雄蕊は通常の機能をもつ。中間にある1個の雄蕊は、両者の中間的な特徴をもつ[39](下図19a)。ショウガ科は1個の機能する雄蕊と2個の仮雄蕊をもち、2個の仮雄蕊は合着して花弁状の構造(唇弁)になっている[40](下図19c)。

_L.,_Sp._Pl._1_273_(1753)._(50611834517).jpg)
.jpg)
これらの仮雄蕊の機能は不明のものが多いが、ツユクサでは仮雄蕊が花弁と共に送粉者への目印として機能していると考えられている[39](上図19a)。またアウストロバイレヤ科では、機能する雄蕊と雌蕊の間にある仮雄蕊が、最初は雄蕊の葯を覆っているが、後に雌蕊を覆うように内側に倒れる[41]。これによって葯が露出し、他家受粉が誘導される。
内部構造
[編集]

花糸はふつう軸状の構造で1本の維管束をもつ(上図5)が、葉状雄蕊では3本の維管束が存在することがある[11]。
葯は1層の表皮で覆われるが、成熟した葯では、表皮細胞は引き延ばされて薄くなっている[18](図20b)。表皮直下に1層から数層の内被(endothecium)が存在し、成熟すると内被の細胞壁はふつう縞状に肥厚する[18](図20b)。葯は、この内被細胞の乾湿運動によって裂開する。さらに内被の内側には数層の細胞からなる中間層(middle layer)が存在する(図20a)が、花粉形成時には崩壊する[18](図20b)。
葯の中で、花粉嚢は1–2層のタペート細胞(tapetal cell)からなるタペート組織(タペータム tapetum)で囲まれている[18][42](図20)。タペート組織は形成中の花粉に栄養を移送し、形成初期につくられた花粉粒(小胞子)の細胞壁(おもにカロースからなる)を分解、花粉粒細胞壁の外層(おもにスポロポレニンからなる)および花粉粘着物(ときに自家不和合性に関わる糖タンパク質を含む)の形成に寄与すると考えられている[18]。葯のタペート組織には、以下の2型が知られている。
- 分泌型(側膜型、腺型; secretory, parietal)[18][42]
- タペート細胞は花粉形成過程を通してもとの位置に留まり、花粉嚢内に物質を分泌し、最終的に崩壊する。
- アメーバ型(侵入型、周辺質型; amoeboid, invasive)[18][42]
- タペート細胞は互いに融合して大きな原形質塊(周辺変形体 periplasmodium)として花粉嚢内に侵入、花粉母細胞や小胞子を取り囲む。この原形質も最終的に崩壊する。
発生
[編集]花を構成する要素は、A, B, C, D, E 遺伝子とよばれるホメオティック遺伝子(原基がどのような器官になるのかを決める調節遺伝子)の産物の組み合わせによって、その分化が制御されている(ABCモデル)[43]。花芽において、葉状の原基にB遺伝子とC遺伝子、E遺伝子が発現することによって、その原基は雄蕊へと分化する。
ギャラリー
[編集]-

-

-

-

-

-

-

-

-

-

-

-

-

-

-
 二体雄蕊
二体雄蕊 -

-
![ヒトリシズカ(センリョウ科)の花序: 1個の花は雌蕊に付着した3個[注 1]の雄蕊からなる](//upload.wikimedia.org/wikipedia/commons/thumb/a/a1/Chloranthus_japonicus_%28flower_s3%29.jpg/120px-Chloranthus_japonicus_%28flower_s3%29.jpg)
-

-

-

-
 二強雄蕊
二強雄蕊 -
 四強雄蕊
四強雄蕊 -

-

-

-

-

-

-

-


.jpg)


_sl14.jpg)


.jpeg)


.jpg)



.jpg)
_pimpollo_2_d%C3%ADas_antes_de_la_antesis,_parte_del_perianto_removida,_anteras_sin_dehiscencia.jpg)
![ヒトリシズカ(センリョウ科)の花序: 1個の花は雌蕊に付着した3個[注 1]の雄蕊からなる](/wiki/%E3%83%95%E3%82%A1%E3%82%A4%E3%83%AB:Chloranthus_japonicus_(flower_s3).jpg)
_Citron_Alba_2zz.jpg)



.jpg)
_sl35.jpg)




_sl13.jpg)

脚注
[編集]注釈
[編集]- ^ または3裂した1個の雄蕊
出典
[編集]- ^ a b (学術用語集 1990, p. 301)
- ^ a b c (学術用語集 1990, p. 292)
- ^ (学術用語集 1990, p. 115)
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao ap aq ar as at au av aw ax ay az ba bb bc bd be bf bg bh 清水建美 (2001). “雄しべ”. 図説 植物用語事典. 八坂書房. pp. 26, 42–56. ISBN 978-4896944792
- ^ a b c d e f g h i j k l m n o p (生物学辞典 2013, pp. 1409–1410)
- ^ a b c d e 長谷部光泰 (2020). “16.6 2胞子嚢を2組もつ雄性胞子葉”. 陸上植物の形態と進化. 裳華房. pp. 223–225. ISBN 978-4785358716
- ^ a b c d e f g (生物学辞典 2013, p. 1403)
- ^ a b c d e f g h i j k l m n o p 原襄, 福田泰二 & 西野栄正 (1986). “おしべ”. 植物観察入門. 八坂書房. pp. 25–26. ISBN 978-4896944792
- ^ a b c d e f 熊沢正夫 (1979). “2) 雄ずい”. 植物器官学. 裳華房. pp. 17−20. ISBN 978-4785358068
- ^ a b 池内昌彦, 伊藤元己, 箸本春樹 & 道上達男 (監訳) (2018). “被子植物の生殖的適応には花と果実がある”. キャンベル生物学 原書11版. 丸善出版. pp. 739–747. ISBN 978-4621302767
- ^ a b c d e f 田村道夫 (1999). “葉的ずいと茎的雄ずい”. 植物の系統. 文一総合出版. pp. 153–157. ISBN 978-4829921265
- ^ a b c アーネスト・ギフォード & エイドリアンス・フォスター (著) 長谷部光泰, 鈴木武 & 植田邦彦 (監訳) (2002). “雄蕊と心皮の系統発生”. 維管束植物の形態と進化. 文一総合出版. pp. 530–540. ISBN 978-4829921609
- ^ Claßen-Bockhoff, R. & Frankenhäuser, H. (2020). “The ‘Male Flower’of Ricinus communis (Euphorbiaceae) interpreted as a multi-flowered unit”. Frontiers in Cell and Developmental Biology 8. doi:10.3389/fcell.2020.00313.
- ^ a b c d 原襄 (1994). 植物形態学. 朝倉書店. p. 152. ISBN 978-4254170863
- ^ (学術用語集(半葯) 1990, p. 60)
- ^ a b c Simpson, M. G. (2019). “Stamens”. In 3rd. Plant Systematics. Academic Press. p. 170. ISBN 978-0128126288
- ^ (学術用語集(花粉嚢、小胞子嚢、葯室) 1990, pp. 102, 287, 292)
- ^ a b c d e f g h i Bhojwani, S.S., Bhatnagar, S.P. (著), 足立 泰二, 丸橋 亘 (訳) (1995). “小胞子嚢”. 植物の発生学. 講談社. pp. 10–23. ISBN 978-4061537088
- ^ (学術用語集 1990, p. 240)
- ^ a b (学術用語集 1990, p. 272)
- ^ Islam, M. S. U., Abid, R. U. B. I. N. A. & Qaiser, M. (2008). “Anther types of dicots within flora of Karachi, Pakistan”. Pak. J. Bot. 40 (1): 33-41.
- ^ (学術用語集 1990, p. 42)
- ^ (学術用語集 1990, p. 239)
- ^ (学術用語集 1990, p. 164)
- ^ (学術用語集 1990, p. 312)
- ^ (学術用語集 1990, p. 132)
- ^ Kabeya, Y. & Hasebe, M.. “被子植物”. 陸上植物の進化. 基礎生物学研究所. 2023年8月12日閲覧。
- ^ Simpson, M. G. (2005). “Androecium”. Plant Systematics. Academic Press. pp. 371–374. ISBN 978-0126444605
- ^ 戸部博 (1992). “ラカンドニア Lacandonia の花(雄しべと雌しべが逆転した花)の意味”. 植物分類,地理 43 (2): 167–168. doi:10.18942/bunruichiri.KJ00001078988.
- ^ (学術用語集 1990, p. 268)
- ^ (学術用語集 1990, p. 171)
- ^ (学術用語集 1990, p. 270)
- ^ (学術用語集 1990, p. 206)
- ^ (学術用語集 1990, p. 51)
- ^ (学術用語集 1990, p. 256)
- ^ (学術用語集 1990, p. 106)
- ^ (学術用語集 1990, p. 309)
- ^ (学術用語集 1990, p. 117)
- ^ a b 多田多恵子 (2002). “ツユクサの用意周到”. したたかな植物たち ―あの手この手のマル秘大作戦. エスシーシー. pp. 54–59. ISBN 978-4886479228
- ^ 「ショウガ科」『日本大百科全書(ニッポニカ)』。コトバンクより2023年8月12日閲覧。
- ^ Kabeya, Y. & Hasebe, M.. “アウストロベイレア目/アウストロベイレア科”. 陸上植物の進化. 基礎生物学研究所. 2021年3月28日閲覧。
- ^ a b c (生物学辞典 2013, p. 878)
- ^ L. テイツ, E. ザイガー, I.M. モーラー & A. マーフィー (編) (2017). “花芽分裂組織と花器官の発生”. 植物生理学・発生学 原著第6版. 講談社. pp. 613–622. ISBN 978-4061538962
参考文献
[編集]- 日本植物学会 (編) (1990). 学術用語集 植物学編 (増訂版). 丸善. ISBN 978-4621035344
- 日本植物学会 (編) (1990). “半葯”. 学術用語集 植物学編 増訂版. 丸善. ISBN 978-4621035344
- 日本植物学会 (編) (1990). “花粉嚢、小胞子嚢、葯室”. 学術用語集 植物学編 増訂版. 丸善. ISBN 978-4621035344
- 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “タペータム”. 岩波 生物学辞典 第5版. 岩波書店. ISBN 978-4000803144
関連項目
[編集]外部リンク
[編集]- 福原達人 (2022) 6-1. 花を構成する要素. 植物形態学. 福岡教育大学. (2022年11月28日閲覧)
- 雄しべに関する記事. 植物Q&A. みんなのひろば. 日本植物生理学会. (2022年11月27日閲覧)